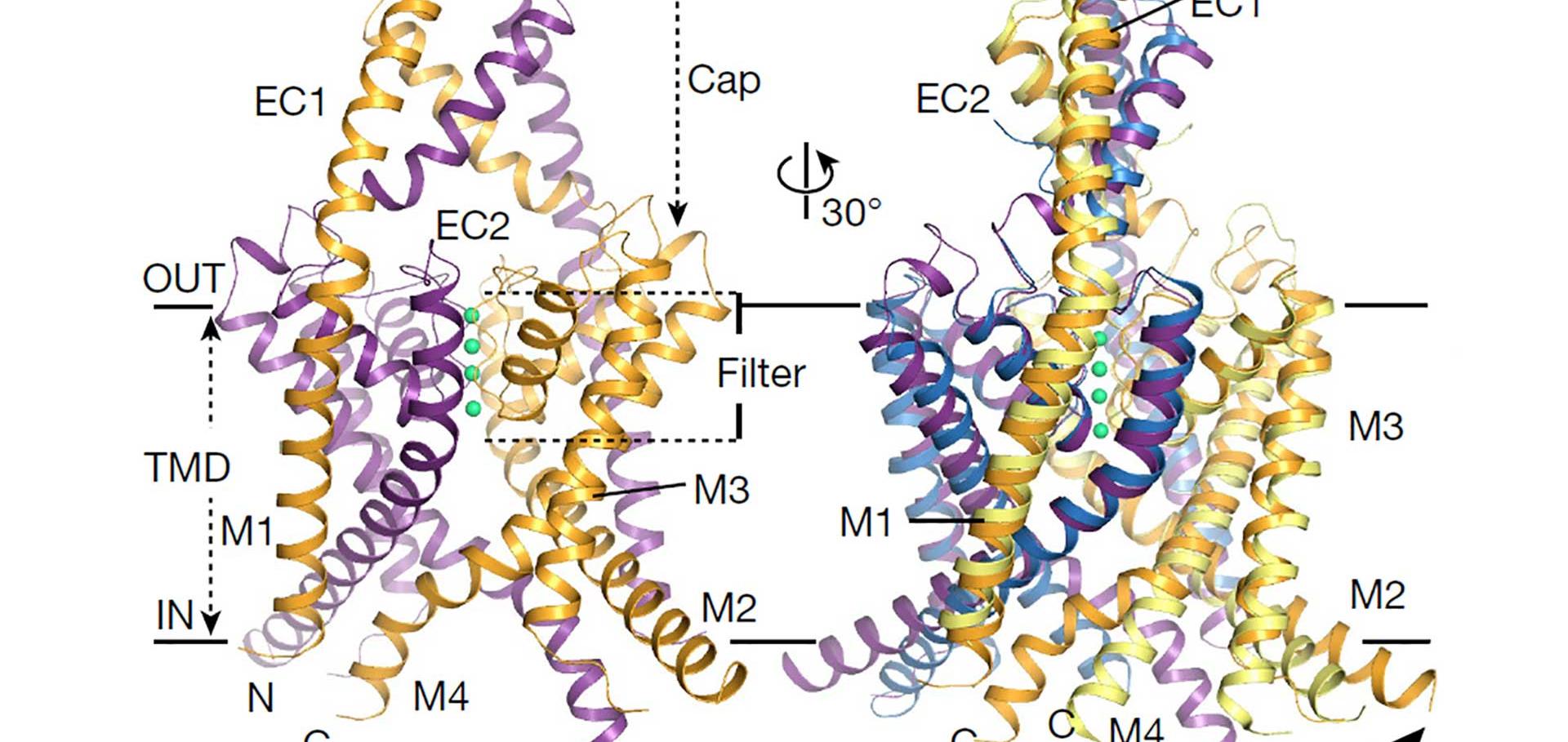
Kir5.1 underlies long-lived subconductance levels in heteromeric Kir4.1/Kir5.1 channels from Xenopus tropicalis.
Biochem Biophys Res Commun 388:3 (2009) 501-505
Abstract:
The inwardly-rectifying potassium channel subunit Kir5.1 selectively co-assembles with members of the Kir4.0 subfamily to form novel pH-sensitive heteromeric channels with unique single channel properties. In this study, we have cloned orthologs of Kir4.1 and Kir5.1 from the genome of the amphibian, Xenopus tropicalis (Xt). Heteromeric XtKir4.1/XtKir5.1 channels exhibit similar macroscopic current properties to rat Kir4.1/Kir5.1 with a faster time-dependent rate of activation. However, single channel analysis of heteromeric XtKir4.1/XtKir5.1 channels reveals that they have markedly different long-lived, multi-level subconductance states. Furthermore, we demonstrate that the XtKir5.1 subunit is responsible for these prominent subconductance levels. These results are consistent with a model in which the slow transitions between sublevel states represent the movement of individual subunits. These novel channels now provide an excellent model system to determine the structural basis of subconductance levels and contribution of heteromeric pore architecture to this process.Contributions of the central hydrophobic residue in the PXP motif of Voltage-Dependent K+ Channels to S6 flexibility and Gating Properties
Biophysical Journal Elsevier 96:3 (2009) 656a
Gating Sensitive Residues In The Pore Of An Inwardly Rectifying Potassium (Kir) Channel
Biophysical Journal Elsevier 96:3 (2009) 462a
State-dependent Cysteine Modification during pH and PIP2 Gating in Kir Channels
Biophysical Journal Elsevier 96:3 (2009) 462a
Contribution of the central hydrophobic residue in the PXP motif of voltage-dependent K+ channels to S6 flexibility and gating properties
CHANNELS 3:1 (2009) 39-45