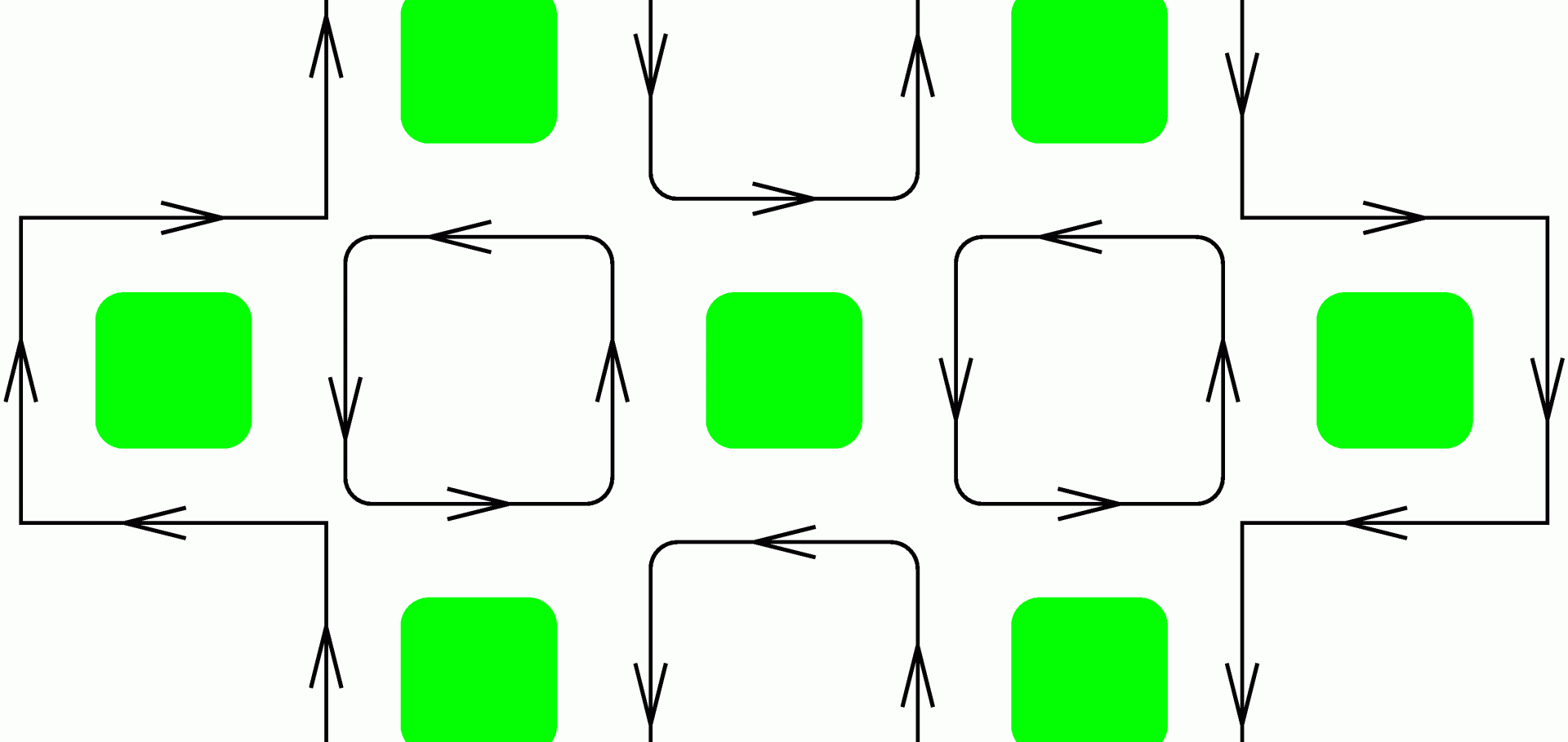
Excitations in the Higher Lattice Gauge Theory Model for Topological Phases III: the (3+1)-Dimensional Case
(2022)
Excitonic fractional quantum Hall hierarchy in Moiré heterostructures
Physical Review B American Physical Society 105:23 (2022) 231521
Abstract:
We consider fractional quantum Hall states in systems where two flat Chern number C=±1 bands are labeled by an approximately conserved 'valley' index and interchanged by time reversal symmetry. At filling factor ν = 1 this setting admits an unusual hierarchy of correlated phases of excitons, neutral particle-hole pair excitations of a fully valley-polarized 'orbital ferromagnet’ parent state where all electrons occupy a single valley. Excitons experience an effective magnetic field due to the Chern numbers of the underlying bands. This obstructs their condensation in favor of a variety of crystalline orders and gapped and gapless liquid states. All these have the same quantized charge Hall response and are electrically incompressible, but differ in their edge structure, orbital magnetization, and hence valley and thermal responses. We explore the relevance of this scenario for moir'e heterostructures of bilayer graphene on a hexagonal boron nitride substrate.Chemotactic self-caging in active emulsions
Proceedings of the National Academy of Sciences of the United States of America Proceedings of the National Academy of Sciences 119:24 (2022) e2122269119
Self-sustained oscillations of active viscoelastic matter
Journal of Physics A: Mathematical and Theoretical IOP Publishing 55:27 (2022) 275601
Abstract:
Models of active nematics in biological systems normally require complexity arising from the hydrodynamics involved at the microscopic level as well as the viscoelastic nature of the system. Here we show that a minimal, space-independent, model based on the temporal alignment of active and polymeric particles provides an avenue to predict and study their coupled dynamics within the framework of dynamical systems. In particular, we examine, using analytical and numerical methods, how such a simple model can display self-sustained oscillations in an activity-driven viscoelastic shear flow.Activity gradients in two- and three-dimensional active nematics
(2022)